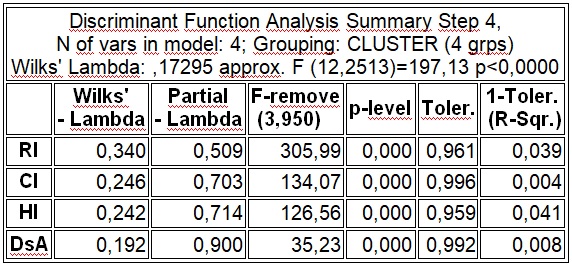
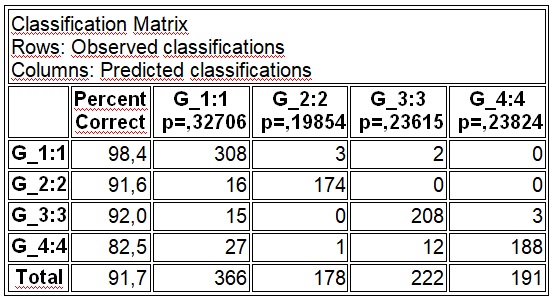
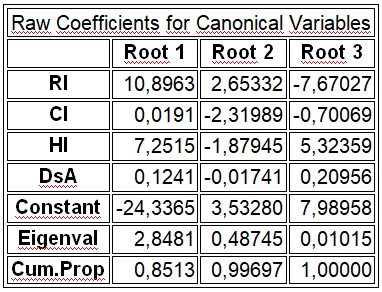
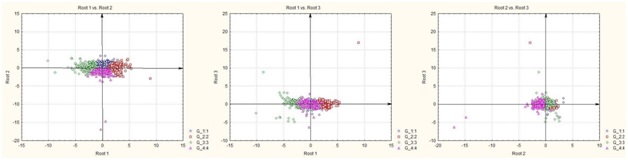
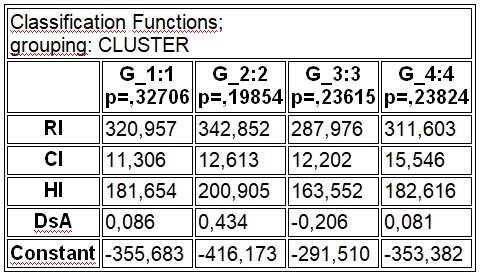
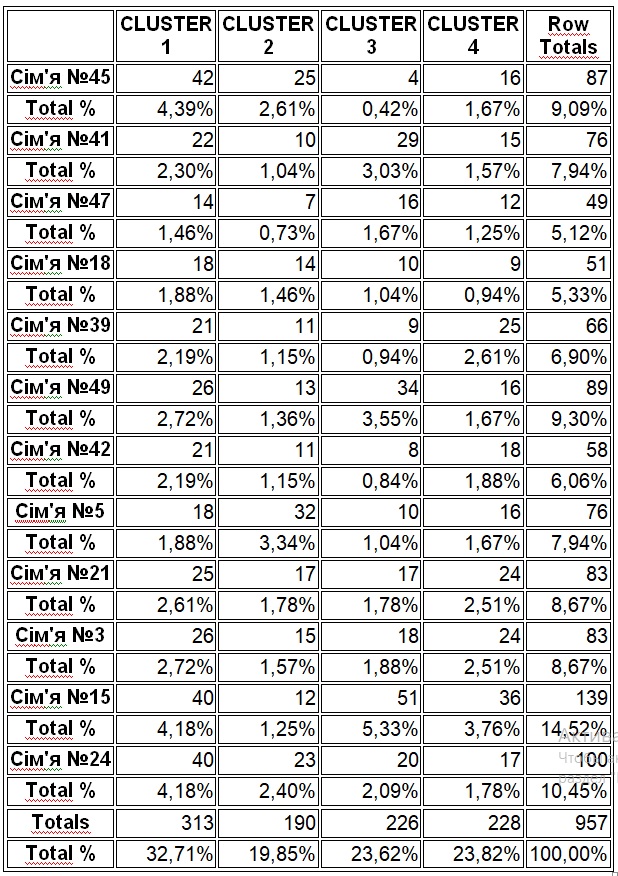
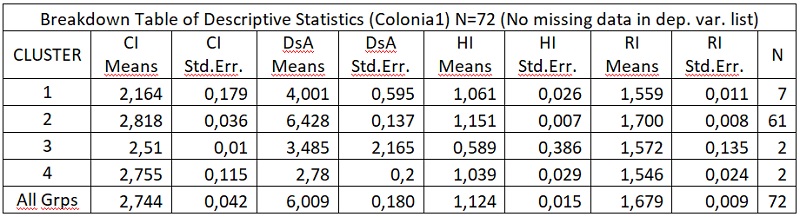
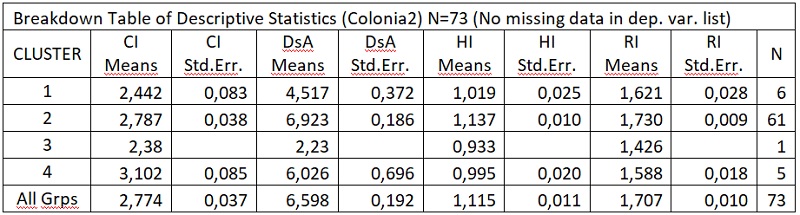
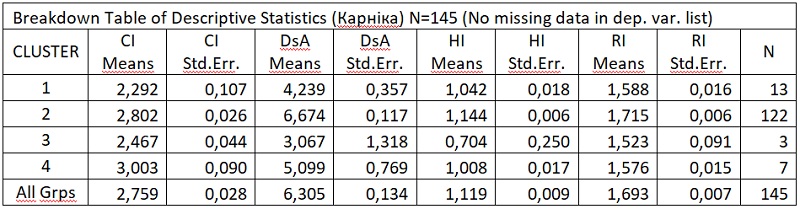
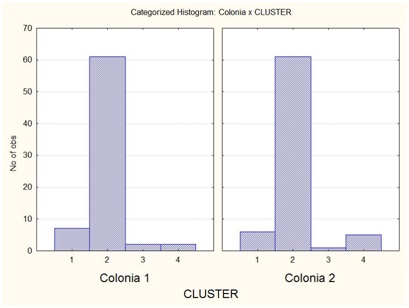
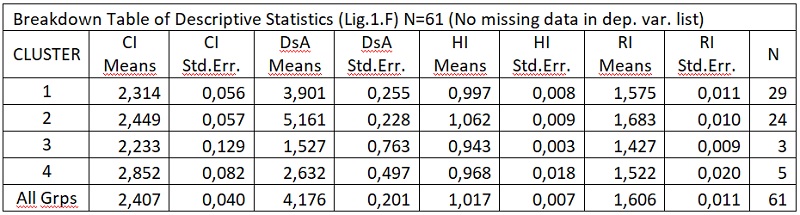
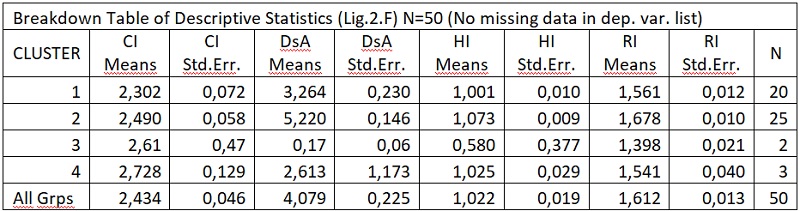
В результате отмеченного и проводившейся безсистемной метизацией в отрасли пчеловодства сложилась довольно тревожная ситуация: исчезли Кубанские пчелы, Крымские, оказались на грани исчезновения пчелы украинской степой породы. В связи с отмеченным, снизилась медопродуктивность отрасли в целом, появились ранее нерегистрируемые болезни пчел.
Член-корреспондент ВАСХНИЛ А.Н.Мельниченко в журнале “Пчеловодство” №9 за 1987 год задает вопрос: “Почему до сих пор ни в одной стране, в том числе в СССР, не созданы селекционные породы медоносной пчелы? И отвечает: “Имеются основания утверждать, что причины этого заключаются, во-первых, в недостаточном знании специалистами пчеловодства современной теории селекции, во-вторых, в ошибочности подмены селекционный теории техническими мероприятиями завоза пчелиных семей и маток южных пород в новые для них природно-климатические области.
Так, разведенческие хозяйства ежегодно вместо выведения методами научной селекции высокопродуктивных пород медоносной пчелы, завозят тысячи пчелиных маток и семей из областей Кавказа, Карпат и даже из зарубежных стран на производственные пасеки в качестве якобы готовых пород… Общим результатом размножения семей всех завозимых южных пород является вымирание большей их части, начиная с третьего-четвертого завозного поколения вследствие близкородственного (инбредного) размножения, а также стихийного скрещивания с местными пчелами и образования на этой основе маложизнеспособных и низкопродуктивных помесей.
…Как массовый, так и единичный завоз пчелиных маток и семей южных пород при отсутствии квалифицированной селекционной работы приводит к разрушению ценных местных популяций, а также к прекращению совершенствования племенного состава пчелиных семей на производственных пасеках. Но, сторонники идеи завоза пчелиных маток в качестве готовых пород, утверждают, что так как якобы освобождают пчеловодов от работы, которую могут более успешно выполнить матковыводные хозяйства.”
В.А. Гайдар, В.П. Полищук, И. И. Головецкий подтверждают: “На породному складі аборигенних бджіл, особливо українських,на жаль,негативно позначилося запозичення сімей та маток іншого походження.В результаті завезення в Україну бджіл сірої гірскої казказьскої та інших порід, місцеві бджоли стали на багатьох пасіках помісями невідомих поколінь. Цей процес став дуже інтенсивним у другій половині XX століття. Починаючи з 1944-1945 років,в Україну для відновлення зруйнованого війною бджільництва масово завозили бджіл переважно кавказького похоження.
Друга хвиля інтродукції бджіл в ареалі української породи після того, як низка наукових установ стала переконувати в ефективності схрещування сірих гірських кавказьких бджіл з місцевими за масштабами завезення була не меншою… Помісні бджоли знизили продуктивність, стали чутливішими до збудників хвороб та інших чинників середовища.(“Визначення породної належності бджіл методом ДНК-тестування, журнал”УП” №1 за 2005 р).
Уважаемые господа-авторы приведенных выше слов, вы говорите о завозе в Украину, в ареал пчел украинской степной породы кавказских пчел и умалчиваете,что в этот ареал в крупномасштабном плане завозились семьи и матки пчел карпатской породы. Вот что пишет бывший директор «Дослідного племінного бджологосподарства” Мукачівське” В.П.Пилипенко: -Так,одним з осередків української степової бджоли називають Кіровоградську область. Але які там можуть бути степові бджоли, коли з 70-х років пасічники різних районів цієї області, в тому числі Знаменського, з зони Карпат (тобто не лише з Закарпаття) завозять тисячі маток і пакетів бджіл. Тисячі пакетів безпосередньо з Закарпаття були туди завезені після 1994 р. При цьому пакети були придбані як любителями, так і господарствами та підприємствами. А через певний час чи не з’явиться десь оголошення,що в Кіровоградській області продають бджіл української степової породи?”. ( журнал “УП”№5 за 1997 р.).
И эта экспансия пчел карпатской породы продолжается не смотря на “План породного .районирования”.
Сейчас уже многие пчеловоды в отличие от некоторых ученых поняли, что пора прекратить работы по смешиванию пород пчел. Вот что пишет, например, М. Матяшев из Черниговской области: “Деякі науковці і практики на догоду регіональним або приватним комерційним інтересам готові повністю знищити українську степову породу, відвівши їй “резервацію”- “одну або декілька областей” їхнього чистопородного розведення і заповнивши решту областей карпатськими бджолами, чим намагаються довести, що нібито – шляхом приливу нової крові карпатських бджіл – можна поліпшити українську степову породу. (журнал “УП”№1 за 2003 р.).
По нашему мнению, ученые должны заниматься наукой, а не коммерцией.
А уж если ради коммерческих интересов растаптываются законы природы и осуществляется ознательное ведение дела к уничтожению аборигенной породы пчел, то это – преступление. Если бы по этой причине лишали таких ученых соответствующих степеней, пчеловодство Украины уже давно было бы рентабельным.
К счастью, наконец поставлены точки над “і”. В.А.Гайдар (институт пчеловодства им. П. И. Прокоповича), В. П. Полищук, И. И. Половецкий (Национальный аграрный университет) в статье “Визначення породної належності бджіл методом ДНК-тестування”, опубликованной в УП”№1 за 2005 р., сообщают: “Відібрані і законсервовані до вимог методики проби бджіл були надіслани для ДНК-тестування (м.Москва). Лабораторні аналізи виконані науковцями А.Б. Абрамовою і О.С. Чудиновим згідно зі спеціальним замовленням… Порівнення результатів досліджень на генетичну дактилоскопію показало не тільки добре виражені породні відмінності українських і карпатських бджіл, а й відсутність у ник ознак сірої гірської кавказької і середньоросійської порід, досліджених раніше в лабораторії “.
Более подробная статья по этому вопросу опубликована в журнале “Пасіка” № 11 за 2007 год. Авторы В.П.Полищук, И.И.Головецкий, В.В,Скрипник сообщают :
“Встановлена в наших дослідах видова специфічність генома бджіл відібраних в типових умовах ареалу їх існузання (Карпати і Лісостеп України), дає підставу стверджувати про належність карпатських та українських бджіл до різних порід.”
Может быть известные ученые, проводившие широкомасштабную интродукцию пчел, в частности, карпатской породы в ареал украинских степных пчел не знали, что нельзя смешивать разные породы? Нет. Не могли не знать.
Еще в 1874 году Ш. Дадан предупреждал, что помеси различных пород нежелательны, так как их качества неустойчивы и меняются с каждым новым поколением.
В 1985 году в журнале”Пчеловодство”/Россия/ была опубликована статья главного редактора журнала “Пчела”/ФРГ/ доктора К. Дреера. Приводим отдельные выдержки из этой статьи: “Хотя каждая порода обладает определенной широтой изменчивости, но ее границы наследственно закреплены и не могут быть нарушены. Так, пчелы, происходящие из жарких стран, гибнут или влачат жалкое существование в холодных районах, и наоборот.
В эти естественные структуры грубо вторгаются люди-селекционеры и пчеловоды-практики,что может привести к уничтожению старых, сложившихся в течение веков пород… В обширном регионе самым приемлемым и перспективным следует признать разведение только одной породы пчел с отбором семей по продуктивности и контролем по породным признакам для выявления возможного появления помесей… В настоящее время исходные породы медоносной пчелы в Европе и за ее пределами оказались генетически разрушенными или разрушаются вследствие беспорядочного скрещивания… Невосполнимая наследственная основа, которая создавалась в течение долгой эволюции, находится сейчас под угрозой гибели… Об этой опасности известно давно,поэтому уже в 1967 году на Международном конгрессе Апимондии в Мэрилэнде (США) была принята рекомендация создавать резерваты для эндемических пород пчел… … Разрушение наших пчелиных пород идет все возрастающими темпами, так что говорить о сохранении некоторых из них, возможно уже поздно… Поэтому я еще раз обращаюсь к организациям пчеловодов и к ученым всего мира, чтобы они, не теряя ни минуты, сделали все для того,чтобы остановить эту разрушительную деятельность.”
К сожалению,наши ученые остались глухи к этому патетическому призыву.Но, мы полагаем, что недалеко то время, когда имена виновников этой “разрушительной деятельности” будут названы.
А что происходит у нас в Украине, в частности, с пчелами украинской степной породы? Приводим краткую историю этого вопроса.
Так, в книге “Селекция пчел”( Г.Д.Билаш, Н.И.Кривцов, 1991 г,) мы читаем: “По происхождению они (украинские степные пчелы, – Р.Е.) представляют южную ветвь среднерусской породы (А. С. Скориков), испытавшую определенное влияние с запада карпатской породы (В.А,Рубин)… Без возражения можно принять только одну гипотезу, суть которой заключается в том,что украинская порода есть не что иное,как результат безсистемной, метизации среднерусской и карпатской пород, а также длительного естественного отбора в степях южной Украины”… О.Н.Полейчук (1913г.) говорил о том,что украинская пчела тождественна краинской во всех отношениях, за исключением зимостойкости, которая в большей мере присуща украинской пчеле”.
В журнале “УП” № 8 за 2001 год В.П.Пилипенко уже утверждает: “0тже, належачи до одного підвиду Apis mellifera carnika, українські і карпатські бджоли можуть бути вихідним матеріалом для отримання внурішньо – породних помісей…Саме прилиття крові чистопородних карпатських бджіл дасть змогу з
часом відновити українську бджолу”.
Но, разве могли все эти гипотезы служить основанием для смешивания разных пород пчел? Нет,не могли. Еще в 2003 году А.Д.Комиссар в журнале “УП” №8 писал: “Докази про те, що ці бджоли (карпатські і українські, -Р.Е.) близьки родичі, практично відсутні”. А А.А. Плахова (Новосибирский государственный университет) отмечает: “Самый быстрый способ уничтожения породы – это массовое скрещивание. Так была уничтожена Крымская порода и неизвестно сколько и где осталось чистопородных среднерусских боровок (журнал “Пчеловодство” №4 за 2003 г.)
Согласно известной заповеди Гиппократа: – “Тот, кто делает успехи в науках и отстает в морали, тот больше вредит, нежели приносит пользу”.
В.А.Губин в 1987 году писал: “Научные работники, делающие на основе многолетних исследований вывод о целесообразности использованя тех или других пчел, должны не только это понимать, но и нести ответсвенность за доброкачественность рекомендаций, от которых зависит дальнейшее состояние как отдельных пасек, так и отрасли в целом”.(журнал “Пче-ловодство” №7 за 1987 г).
В.П.Куринной писал: “Академик Т.Д.Лисенко вважаючи себе знавцем, отримавши владу,на десятирічча загальмував розвиток генетики,что равноцінно злочину”.
Да, “злочину” и “злочину” также в том,что по его вине академик Н.И. Вавилов умер в тюремном застенке.
Несомненно, пчеловодству нанесен огромный вред и не только в странах СНГ,но и в западных государствах.
В. А. Рубин писал:”Ушедшее столетие отмечено в истории пчеловодства как век неожиданностей, приведших к почти полному исчезновению чистопородных пчел. Теперь на пасеках царит генетическая неразбериха (ж.”Пчеловодство” №1 за 2001 год). ;
Следствием смешивания пород пчел является прежде всего потеря пчелами иммунитета на генном уровне, в результате чего наши пчелы оказываются незащищеными по отношению к заболеваниям, которые ранее вообще не наблюдались в пчеловодстве.
Так,профессор 0.Ф. Гробов (Всероссийский институт экспериментальной ветеринарии) отмечает: “В результате проведенных работ установлены новые нозологические единицы ранее нерегистрируемых в стране (в СССР, – Р.Е.) заболеваний, таких как спироплазмоз, гафниоз, колибактериоз, протеозы, шигеллез, цитобактероз” журнал”Пчеловодство” №2 за 199б г.).
Было бы неверным утверждать,что болезнетворных бактерий ранее не было в зоне обитания пчел. Были,конечно. Но пчелы были невосприимчивы к этим болезням. А, значит, иммуная система пчел перестала обеспечивать их здоровье от этих заболеваний.
Что же могло ослабить, кроме отмеченного, иммунитет пчел, его защитные свойства? Причин здесь много. Прежде всего иммуная система не успевает или, по крайней мере, запаздывает в слежении за изменяющейся экологической обстановкой. Появление каких-либо новых болезней, таких как варроатоз, аскосфероз и т.д. приводят к ослаблению защитных свойств организма и провоцирует восприимчивость к другим заболеваниям, которые ранее не наблюдались. (Источником изложенного являются статьи, опубликованные в пчеловодных журналах).
Несомненно, серьезное значение имеет перемещение пчел человеком из одной экологической среды в другую со всеми их болезнями, безсистемная метизация.
Как следует из сказанного ранее,в каждом ареале должна разводиться лишь одна порода пчел и, именно та, которая является аборигенной, т.е. порода, которая испокон веков обитала и эволюционировала в данной зоне одновременно с изменяющимися внешними условиями. Такой породой в Украине, исключая зоны Карпат и Полесья, являются пчелы украинской степной породы. И нет никаких шансов, что если этих пчел завести, например, на Камчатку, их организм не будет протестовать и что это в конечном счете не скажется на их здоровье, на медопродуктивности, на зимостойкости и прочих характеристиках. И отвественные ученые ни в коем случае такого переселения рекомендовать не могут.
Может быть есть породы пчел, которые значительно превосходят украинских пчел по медопродуктивности? Может быть они недостаточно устойчивы к болезням? Может быть они недостаточно зимостойки? Может быть они злобливы, что с ними трудно работать? Может быть, они нечистоплотны, не борются с молью и у них планки рамок продырявлены личинками моли? Может быть они слишком ройливы? Может быть они воровиты? Нет,нет и нет!!!
Эти пчелы так дороги для Украины,что нет им цены. Их надо беречь, как бесценное богатство, которым наградил Бог нашу Украину!!!
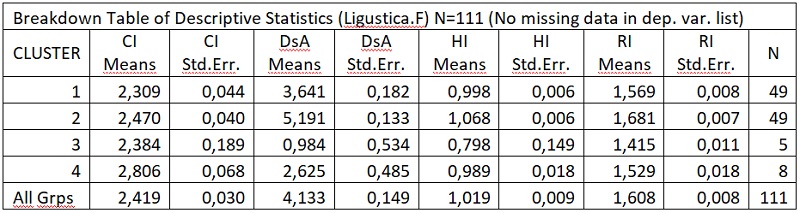
А теперь приводим основные характеристики этой украинской породы пчел согласно данным, приведеным в журнале “Пасіка”№11 за 2007 г., в статье В.П.Полищука, И.И.Головецкого, В.В.Скрипника “Селекция украинских пчел”:
Довжина хоботка – 6,34-6,63 /в середньому 6,50 мм/;
Кубітальний індекс – 2,18-2,62;
Дискоїдальне зміщення позитивне у 72-94% бджіл;
Форма заднього краю воскового дзеркальця п’ятого стерніта – випукла у 64-96% бджіл;
Окраска бджіл – світлосірі;
Агресивність – помірно агресивні, не схильні до нападу і крадіжки меду з гнізд сімей і відводків;
Печатка меду в стільниках – світлою восковою кришечкою;
Зимостійкість – зимують добре;
Стійкість до проносих захворювань – підвищена;
Розвиваються весною – інтенсивно;
Схильність до роіння – помірна; Пристосованість до збору нектару під час посередніх і сильних взятків – добра;
плодючість маток – 1950-2300 яєць за добу с забезпеченням нарощуваннясильних сімей до медосбору з білої акації, гречки, липи та інших медоносів, та наприкінці літа при підготовці до зимівлі;
За даними І.0. Левченка бджоли української породи починають здійснювати сигнальні рухи при вмісті цукру в розчині 18%. Цей показник є значно вищим, ніж у бджіл карпатської породи (8%), що може бути контрастною ознакою при визначанні породної належності бджіл. В умовах задовільних і багатих ресурсів нектару збір меду за сезон становить 60-80 кг на сім’ю, а за сприятливої погоди – понад 105 кг.(ж.”УП”№1 за 2007 р).
“За сучасних умов збіднених ресурсів нектару бджолині сім’ї (Української степової породи, – Р.Е.), що збереглися в чистоті породи, збирають по 70-80 кг, а в окремі роки – понад 120 кг. (“В.П.Поліщук,”Бджільництво”,2001).
Приводим сводку отзывов о пчелах украинской степной породы известных пчеловодов недалекого прошлого и настоящего времени.
Так П.Л.Снежневский о достоинствах “украинок”еще в 1910 году сообщал,что в условиях Воронежской губернии украинские пчелы по медопродуктивности превосходили среднерусских пчел. Он обратил внимание на их дальнелётность, выносливость, спокойное поведение при осмотре гнезда,миролюбие, малую склонность к роению; хорошую зимостойкость, красивую печатку меда, незначительное прополисование гнезда и называл их “нашей украинской жемчужиной” и, изучив их, не расставался с ними до конца своей жизни». В 1913 году П.Л.Снежневский пытался привлечь внимание к украинским пчелам заграничных коллег, однако с горечью отмечал,что “…Сами же мы,очевидно не оценим своего пчелиного сокровища”.Так и произошло.
Известные в прошлом пчеловоды-промышленники В.Ф.Ващенко, Ю.С.Шелухин имели крупные пасеки украинских степных пчел, получая от них высокие доходы. Хорошо отзывались об украинских пчелах О.Н.Полейчук, писавший и том, что украинские пчелы по зимостойкости и плодовитости маток превосходят краинок; И.Федин, отмечавший такие важные свойства этих пчел, как миролюбие, высокие строительные качества и несильную ройливость./1913 – 1924 гг/.
Высокую оценку украинским пчелам дал В.В.Алпатов /1948/ ,отметив их большую пригодность для опыления клевера, он приводит целую сводку отзывов разных авторов об этих пчелах, как “кротких”, трудолюбивых и доходных”.
По мнению В.А.Нестерводского (1966г.) украинская пчела продуктивна, энергична,трудолюбива и малоройлива.Она очень острожно покидает гнездо, поэтому ее мало гибнет в холодную погоду. Расплод у нее сплошной; семьи быстро набирают силу.
Важно отметить,что согласно оценкам Г.Ф.Таранова (1982г.) украинские пчелы устойчивы к заболеваниям.
Г.Д. Билаш отмечает: “Зарегистрированы случаи, когда украинские степные пчелы в условиях сильного устойчивого медосбрра собирали до 100 кг меда в среднем по пасеке, (“Селекция пчел”,1991г.).
Следует сказать, что более ста лет назад эти пчелы были завезены украинскими переселенцами на Дальний восток, где они до настоящего времени сохранились в относительной чистоте – имеются, сведения, что при медосборе с липы там они приносят до 32 кг нектара в день и общая продуктивность донтигает 400 кг меда. Исходя из этого,можно полагать,что здесь на родине они обладают большим медопродуктовым потенциалом.
В книге “Пчеловодство.Маленькая энциклопедия”, М.2000, Научное издательство “Большая Российская энциклопедия”,мы читаем:”Степные украинские пчелы – порода, сформировавшаяся в условиях степи и лесостепи Украины. Хорошо приспособлена к морозной зиме и жаркому, относительно сухому лету… Степные украинские пчелы значительно легче, чем среднерусские, переключаются из роевого состояния в рабочее; для этого иногда достаточно вырезать все роевые маточники и расширить гнездо.Тихая смена маток наблюдается довольно часто.Зимостойкость и устойчивость к нозематозу, европейскому гнильцу, и особенно – падевому токсикозу, высокие. Степные украинские пчелы миролюбивее среднерусских; при осмотре гнезда ведут себя на сотах, вынутых из улья, относительно спокойно. Умеренно прополисуют гнездо. По предприимчивости в отыскании новых источников корма и скорости переключения с худших на лучшие, они лишь немного превосходят среднерусских и уступают в этом отношении серым горным кавказским пчелам… Приспособлены к сбору нектара со степного разнотравья. Печатка меда преимущественно белая (сухая). Медовая продуктивность пчелиных семей высокая”.
Следует отметить,что многочисленные данные по сравнительному изучению украинской и серой горной кавказской пород (П.А»Губа,1965 г.) в местностях и в годы со скудными условиями медосбора показывают,что “украинки” имеют преимущество над кавказянками. Характерно,что украинские пчелы вкладывают мед сначала в магазинную часть гнезда и затем уже в расплодную, т.е. ограничение яйцекладки маткой наступает после заполнения свободной части улья. В период же сильного медосбора пчелы “заливают” и гнездовую часть улья, заполняя все свободные ячейки и значительно ограничивают работу матки.
При содержании сахара в нектаре менее 18% пчелы из улья не вылетают и берегут свои резервы. Известно,что краинские пчелы летят за нектаром при достижении сахаристости 8%. Не трудно определить, что для получения из такого нектара 10 кг меда пчелы должны принести в улей 100 кг и затем 90 литров воды выпарить. Для получения 10 кг меда с 18%- ным содержанием сахара в нектаре украинским пчелам надо принести в улей вдвое меньше нектара, точнее – 45,5 кг и выпарить лишь 35,5 кг воды». Из этого можно сделать вывод,что при минимальной, граничности содержания сахара в нектаре, работа украинских степных пчел в два с лишним раза эффективнее краинок.
Дальнелетность пчел разных пород изучалась И.А.Левченко (Институт зоологии АН УССР). По его данным украинские степные пчелы как и среднее русские имеют привязанность к найденному сильному источнику нектара и, как правило (при размещении пасеки,например,в центре массива медоносов), работают преимущественно на расстоянии до 500 метров и около половины пчел рассеиваются не далее 1,5 км от точка. Однако, при отсутствии взятка вблизи пасеки пчелы могут летать на расстояние до 5 км.
Еще один важный аспект. Известно,что переносчиком болезней является, в частности, восковая моль. Как отмечает Н.И.Столбов (ВНИИ ветеринарной энтомологии, г.Тюмень), кроме непосредственного вреда – порчи сотов, личинки моли механически переносят возбудителей многих болезней. Исследованиями, проведенными Е.Н.Скрыпником и Е.И.Корщонским (Крымская научно-исследоватеяьская станция,1970) устаневлено, что моль является одним из источников заражения пчел гнильцами и другими болезнями расплода пчел. Если в улье, на дне, имеет место слой восковой крошки, – это первое пристанище моли. Мы,работая ранее на государственных пасеках с пчелами завозных пород, убеждались, что все планки рамок в таких семьях продырявлены личинками моли. Мы убеждались так же в том,что в семьях таких пчел всегда имеет место потенциал заражения их гнильцами, аскосферозом и другими болезнями. В семьях украинских степных пчел ничего подобного не наблюдается. Как правило, украинские пчелы исключительно чистоплотны. Весной при первых облетах они незамедлительно приступают к уборке подмора и восковой крошки.
Считаем необходимым отметить,что в результате проведенной нами селекционной работы по очистке генотипа пчел от постороннего “загрязнения” в 1998 году на пасеке основных семей нашего пчелохозяйства без какого-либо лечения и профилактики полностью исчез аскосфероз, хотя в отдельных нуклеусных семейках матковыводного питомника еще наблюдались вспышки этого заболевания после отбора матки.
В последующие годы до настоящего времени ни аскосфероза, ни других болезней, кроме варроатоза в хозяйстве нет.
Известно также,что болезни передаются через пчелиное воровство (И. А.Шабаршов,1988). Украинские степные пчелы неворовиты. Этот недостаток присущ в основном пчелам южных пород. Ульи нашей пасеки весь рабочий сезон находятся на передвижных пасечных установках с расстоянием между ними 17 см с нижними летками высотой 23 мм во всю ширину передней стенки улья. Как правило, пчелиного воровства на пасеке не наблюдается. В 30 метрах от основных семей матковыводного хозяйства расположено более 1000 семеек нуклеусного парка. И здесь при использовании пчелами семеек донного летка обворовывание их не происходит.
Следует отметить еще одну особенность украинских пчел. Поздней осенью, в период кормления пчел на зиму и позднее, пчелы почти полностю перекрывают нижний леток размером 375×23 мм прополисом в количестве не менее 70 г на семью. Сбор пчелами такого количества прополиса в летковом проеме также свидетельствует о факторе, обеспечивающем здоровье семей, так как прополис обладает высокой биологической активностью, на защитные свойства которого от вредителей и болезней указывал С.Н.Поправко.
Важным признаком украинских степных пчел является также их хорошая зимостойкость. Весной, открывая ульи, приятно осознавать отсутствие ноземотоза. Все рамки и соты чистые, пчелы бодрые.
Кроме того,украинские пчелы не только медовитые, но и восковитые.В среднем за сезон при стабильном медосборе пчелы отстраивают не менее 20 сотов в переводе на дадановскую рамку,а лучшие семьи – до 25 сотов. При строительстве сотов украинские пчелы как и среднерусские не скрепляют их восковыми перемычками, что мы можем наблюдать у пчел южных пород. При выемке из гнезда сотов в “семьях южанок перемычки разрываются и мед вытекает из разрушенных ячеек. При обратной постановке сота приходится попеременными приближениями сота к сопрягаемому пытаться не давить пчел, осушивающих разрушенные ячейки,что не может не сказаться на производительности труда, в частности, на промышленных пасеках».













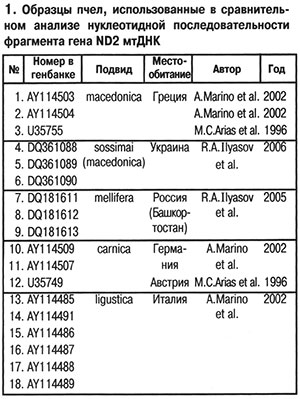 Для уточнения действительного таксономического статуса украинских пчел необходимо было провести дополнительные исследования с использованием молекулярно-генетических методов. С этой целью мы сравнили последовательности нуклеотидов во фрагменте гена ND2 митохондриальной ДНК (мтДНК) у ряда подвидов европейских пчел. Исследование провели на трех образцах пчел с Украины (пасека А.Д.Комиссара около Киева), трех образцов из России (Республика Башкортостан, Бурзянский район: пасеки «Капова Пещера», «Коран-Елга», борти Государственного природного заповедника «Шульган-Таш») и внесли в базы данных международных генетических банков (табл. 1).
Для уточнения действительного таксономического статуса украинских пчел необходимо было провести дополнительные исследования с использованием молекулярно-генетических методов. С этой целью мы сравнили последовательности нуклеотидов во фрагменте гена ND2 митохондриальной ДНК (мтДНК) у ряда подвидов европейских пчел. Исследование провели на трех образцах пчел с Украины (пасека А.Д.Комиссара около Киева), трех образцов из России (Республика Башкортостан, Бурзянский район: пасеки «Капова Пещера», «Коран-Елга», борти Государственного природного заповедника «Шульган-Таш») и внесли в базы данных международных генетических банков (табл. 1).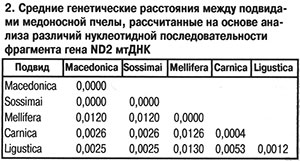 На основании сравнения нуклеотидных последовательностей фрагмента гена ND2 мтДНК пчел с использованием программы MEGA 3.1 (Kumar et al., 2004) и дальнейшего расчета средних генетических расстояний между образцами было показано тесное генетическое родство пчел с Украины (пасека А.Д.Комиссара) и пчел A. m. macedonica из Греции: средние генетические расстояния между ними были равны 0,0000 (табл. 2). Наиболее генетически отдаленными от украинских пчел оказались среднерусские A. m. mellifera из Республики Башкортостан (Россия), что вполне логично, так как A. m. macedonica, A. m. carnica, A. m. ligustica — представители эволюционной ветви C (Ruttner, 1988), а A. m. mellifera — эволюционной ветви М.
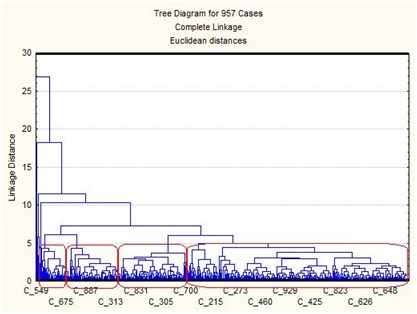
На основании сравнения нуклеотидных последовательностей фрагмента гена ND2 мтДНК пчел с использованием программы MEGA 3.1 (Kumar et al., 2004) и дальнейшего расчета средних генетических расстояний между образцами было показано тесное генетическое родство пчел с Украины (пасека А.Д.Комиссара) и пчел A. m. macedonica из Греции: средние генетические расстояния между ними были равны 0,0000 (табл. 2). Наиболее генетически отдаленными от украинских пчел оказались среднерусские A. m. mellifera из Республики Башкортостан (Россия), что вполне логично, так как A. m. macedonica, A. m. carnica, A. m. ligustica — представители эволюционной ветви C (Ruttner, 1988), а A. m. mellifera — эволюционной ветви М.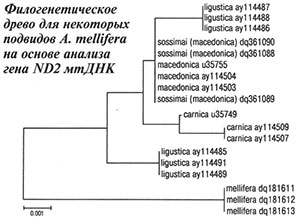 По данным генетических дистанций методом кластеризации ближайшего соседа (Saitou, Nei, 1987) мы построили филогенетическую схему (рис.). На ней видно, что все образцы A. m. sossimai и A. m. macedonica объединились в одну группу, что говорит о тесном генетическом родстве украинских и греческих пчел. В другую группу объединились пчелы A. m. carnica из Германии и Австрии, которые из всех сравниваемых подвидов были наиболее близки к группе A. m. macedonica. Представители A. m. mellifera из Республики Башкортостан оказались наиболее отдаленными от группы, объединяющей A. m. macedonica и A. m. sossimai, а также от групп других подвидов эволюционной ветви С. Образцы пчел A. m. ligustica из Италии разделились на две группы, одна из которых была близка к группе A. m. macedonica, а другая находилась на значительном отдалении от A. m. macedonica и A. m. carnica и была близка к A. m. mellifera. Согласно результатам исследований морфометрических характеристик пчелы из Италии обладали бóльшим сходством с A. m. carnica из Австрии и Югославии (Ruttner, 1988). Возможно, некоторые популяции A. m. ligustica из Италии пересекались с пчелами A. m. mellifera из Франции, что подтверждает полиморфизм межгенного локуса COI-COII мтДНК (Franck et al., 2000).
По данным генетических дистанций методом кластеризации ближайшего соседа (Saitou, Nei, 1987) мы построили филогенетическую схему (рис.). На ней видно, что все образцы A. m. sossimai и A. m. macedonica объединились в одну группу, что говорит о тесном генетическом родстве украинских и греческих пчел. В другую группу объединились пчелы A. m. carnica из Германии и Австрии, которые из всех сравниваемых подвидов были наиболее близки к группе A. m. macedonica. Представители A. m. mellifera из Республики Башкортостан оказались наиболее отдаленными от группы, объединяющей A. m. macedonica и A. m. sossimai, а также от групп других подвидов эволюционной ветви С. Образцы пчел A. m. ligustica из Италии разделились на две группы, одна из которых была близка к группе A. m. macedonica, а другая находилась на значительном отдалении от A. m. macedonica и A. m. carnica и была близка к A. m. mellifera. Согласно результатам исследований морфометрических характеристик пчелы из Италии обладали бóльшим сходством с A. m. carnica из Австрии и Югославии (Ruttner, 1988). Возможно, некоторые популяции A. m. ligustica из Италии пересекались с пчелами A. m. mellifera из Франции, что подтверждает полиморфизм межгенного локуса COI-COII мтДНК (Franck et al., 2000).