Наведено результати вивчення породної належності сімей українських та карпатських бджіл методом ДНК-тестування. Встановлено, що вони належать до різних порід і характеризуються чітко вираженими ознаками геному
Бджола, материнська сім’я, порода, внутрішньопородний тип, генеалогічна група, екстер’єр, ДНК-тестування.
Родючі ґрунти України, багата медоносна рослинність і відповідні кліматичні умови сприяли розмноженню бджіл і збиранню великої кількості меду. Протягом тривалого еволюційного розвитку відбувався природний процес створення аборигенних бджіл, які добре пристосовувались до місцевих умов клімату й медозбору [9]. На території України сформувались і нині районовані українська степова і карпатська породи, а також поліська популяція середньоросійських бджіл [10].
Українські бджоли є найпоширенішою породою в нашій країні і належать до групи аборигенних бджіл європейського походження, займаючи Лісостеп і степову зону. Їхній ареал на Заході охоплює частину прикордонних районів Молдови і доходить до Карпат. На Півночі українська бджола межує з поліською популяцією середньоросійської породи.
Східна частина ареалу українських бджіл представлена рядом сусідніх з Україною районів Російської Федерації. В другій половині ХІХ століття відбулося масове завезення їх з природного ареалу на широкі простори Приморського краю і губерній Східного Сибіру Російської імперії [2]. Як свідчать наукові дані, бджоли українського походження в багатьох районах зберегли свої породні особливості і в умовах багатої медоносної бази дають рекордні збори меду.
Генетичний потенціал української бджоли багатий. За словами В. А. Нестерводського [8], наша бджола гарна й варта уваги, за властивості її хвалять навіть чужі люди. Обстеживши пасіки на Поділлі, Уманщині, Харківщині, він зазначає, що українська бджола жвава, енергійна, працьовита, мало роїться порівняно з південною, далеко літає за поживою і тому продуктивніша.
Вона дуже обережно вилітає, то ж її мало втрачається й сім’ї не знесилюються в прохолодну пору. Вощину будує гарно, мед закриває рівною, білою мов сніг покришкою, тому стільниковий мед має чудовий вигляд. Матки дуже плодючі й розплід у них безперервний, то ж сім’ї швидко стають сильними і відпускають ранні рої.
Карпатські бджоли природно займають гірські та передгірні райони Західної України. До найбільш типових представників, що різняться рядом цінних біологічних особливостей і господарсько корисних ознак, належать бджоли високогірних районів Закарпаття, які пристосовані до умов зимівлі з тривалим безобльотним періодом.
За екстер’єром вони близькі до крайнських бджіл, але відрізняються від них високою зимостійкістю, меншою враженістю нозематозом та іншими етологічними ознаками [4]. Тому карпатських бджіл почали завозити в інші області України. Започатковане Г. А. Аветисяном [3] поглиблене вивчення й селекція карпатських бджіл завершилося створенням в породі популяцій, екотипів, ліній.
На українську породу бджіл, на жаль, негативно вплинули сім’ї та матки інших порід. Починаючи з 1944-1945 рр., в Україну для відновлення зруйнованого війною бджільництва масово завозили бджіл з Кавказу та іншого походження, у тому числі й сірої гірської кавказької породи. На багатьох пасіках відбулось схрещування місцевих бджіл і виникли помісі різних поколінь.
Як масове, так і поодиноке завезення бджолиних маток і сімей інших порід за низького рівня селекційної роботи призвело до знищення цінних місцевих популяцій. Помісні бджоли знизили продуктивність, стали більш чутливими до збудників хвороб та інших чинників середовища. В цих умовах збільшується значення систематичної племінної роботи з районованими породами бджіл як на товарних пасіках, так і в спеціалізованих племінних господарствах.
Для збереження місцевих порід бджіл на пасіках України необхідно застосовувати чистопородне розведення. Важливе значення в цьому має добір селекційного матеріалу, організація репродукції чистопородних бджіл, створення внутрішньопородних типів.
Нині належність бджіл до тієї чи іншої породи визначають переважно за фенотиповими ознаками (екстер’єрними, фізіологічними, етологічними) та показниками господарської цінності. Вони широко варіюють навіть в межах однієї породи або популяції, оскільки залежать від географічного розміщення бджіл, їх віку і живлення [2]. Крім того, велику роль у визначенні ознак породи відіграє кваліфікація спеціаліста.
Тому виникла гостра необхідність в розробці певних заходів на встановлення і збереження біологічної чистоти порід бджіл. Це вкрай важливо як для практичної генетико-селекційної роботи, так і для генетико-популяційних і філогенетичних досліджень.
У 1989 р. О. В. Капралова, Л. В. Дубова [7] розробили цитогенетичний метод вивчення ДНК в ядрах клітин гемолімфи різних стаз бджіл. Як показали результати досліджень, існує достовірна різниця за особливостями ДНК в ядрах клітин гемолімфи різних порід бджіл на деяких етапах онтогенезу. На основі отриманих результатів було запропоновано в практичній роботі для характеристики породи поряд з загальноприйнятою методикою морфометрії користуватись цитофотометричним методом дослідження породної належності бджіл.
Геном бджіл залишається мало дослідженим, побудована лише приблизна генетична карта [12]. Все це свідчить про актуальність вивчення генотипу різних порід бджіл з використанням методів молекулярної генетики.
Одним з прогресивних, порівняно швидких і менш трудомістких є RAPD-аналіз (random amplified polymorphic DNA) [13] генетичного поліморфізму бджіл. Це модифікований метод полімеразної ланцюгової реакції з використанням коротких фрагментів ДНК, що генетично неспецифічні досліджуваній ДНК.
Суть методу полягає в тому, що в ролі праймера (послідовність ДНК з 8-12 пар нуклеотидів) використовується довільний нуклеотид, здатний гібридизуватись з досліджуваною ДНК і ампліфікувати (збільшувати копії) її [5].
RAPD-технологія використовується при вивченні геному, популяцій, типів, маркуванні ознак [6]. У бджільництві ДНК-тестування недостатньо розроблене і використовується мало. Стосовно українських та карпатських бджіл цей метод ідентифікації порід не застосовували.
Мета досліджень — провести ДНК-тестування бджіл української і карпатської порід на чистопородних сім’ях, відібраних за екстер’єрними та іншими ознаками.
У дослідах використали проби бджіл із сімей, що належать до племінного матеріалу, створеного в результаті багаторічної селекційної роботи з карпатською та українською породами.
Досліджувані бджоли української породи належать до хмельницького внутрішньопородного типу, створеного кафедрою бджільництва ім. В. А. Нестерводського (плембджолорозплідник “Прибузькі медобори”, с. Головченці Хмельницької області). Зразки бджіл карпатської породи відібрані з відселекціонованих сімей пасіки відділу селекції і репродукції бджіл карпатської породи Інституту бджільництва ім. П. І. Прокоповича УААН (с. Вучкове Закарпатської області).
Бджоли сімей хмельницького внутрішньопородного типу за екстер’єрними ознаками повністю відповідають вимогам української породи — довжина хоботка у них у середньому дорівнює 6,41 мм, показники кубітального індексу — 2,27, дискоїдального зміщення — позитивне (79%), випукла форма заднього краю воскового дзеркальця п’ятого стерніта спостерігається у 90% бджіл.
Ці дані є результатом комп’ютерної обробки промірів екстер’єру бджіл близько 400 сімей, досліджених за останні п’ять років на завершальному етапі створення типу. Він структурований на дев’ять генеалогічних груп, кожна з яких являє сукупність сімей спільного походження з матками різних поколінь, що є нащадками відібраних родоначальниць.
Відібрані і законсервовані згідно з вимогами методики проби бджіл були надіслані для ДНК-тестування (м. Москва). Лабораторні аналізи виконані А. Б. Абрамовою і О. С. Чудіновим за спеціальним замовленням.
Оскільки справжня назва досліджуваних порід є дискусійною, бджіл української породи з піддослідних сімей хмельницького внутрішньопородного типу умовно позначили як українські степові (Apis mellifera acervorum), а відібрані з сімей пасіки відділу селекції і репродукції карпатських бджіл Інституту бджільництва ім. П. І. Прокоповича — карпатські (Apis mellifera carpatica).
Аналіз проводили за методом RAPD-праймерів — локальної ампліфікації набору фрагментів ДНК розміром від 50 до 2500 пар нуклеотидів, франкованих короткими інвертованими повторами, які виступають як ділянки випалювання для випадково підібраного 10-нуклеотидного праймера. Розподіл у геномі інвертованих повторів унікальний, що дозволяє виявляти відмінності в RAPD-фінгерпринтах різних видів і порід.
Для диференціації порід бджіл підібрано породоспецифічні маркери. В результаті RAPD-ампліфікації утворюється набір фрагментів ДНК різної маси, який при розподілі на форезі дає унікальний фінгерпринт, де кожний поліморфний між видами або внутрівидовий фрагмент ДНК проявляється в ролі видоспецифічного або породоспецифічного маркера. Тому підбір маркерів, які диференціюють породи в RAPD-аналізі полягає у виявленні ефективних праймерів довжиною 10 нуклеотидів.
Для проведення RAPD-аналізу використовувалась тДНК-зразків робочих бджіл, призначених для досліджень. Виділення тотальної ДНК проводилось методом SDS-лізису з наступною фенол-хлороформною екстракцією. Після виділення і очищення ДНК-зразків визначали концентрацію, ступінь чистоти і ступінь фрагментації ДНК.
Ступінь фрагментації тДНК-зразків робочих бджіл проводили методом електрофорезу в 0,8%-ному агарному гелі. Ступінь чистоти і концентрацію препарату тДНК-зразків визначали за допомогою вимірювання оптичного поглинання препаратів тДНК на довжині хвиль 260 і 280 нм.
Локальна ампліфікація при полімеразно-ланцюговій реакції з використанням RAPD-праймерів проводилась згідно з методикою, запропонованою для вивчення генетичного поліморфізму ДНК-генома різних порід бджіл [11]. У дослідженнях використані синтетичні 10-членні олігонуклеотиди, послідовність яких відповідала праймерам, розробленим компанією “Operon” (США). ПЦР-продукти розділяли в 6%-ному поліакриламідному (ПААГ) гелі. Після проведення електрофорезу ПААГ або агарний гель зафарбовували бромистим етидієм. Для візуалізації картини розподілу фракцій ДНК після RAPD-ампліфікації використовували УФ-трансілюмінатор.
Фотовідбиток з результатів дослідів отримували за допомогою гель-документуючої системи “Kodak”.
Результати досліджень. На рисунку представлено результат електрофоретичного розподілу фракцій ПЦР-продукту зразків досліджених бджіл після локальної ампліфікації з використанням праймера В15. Праймер В15, як і випробуваний в дослідах В06, виявився ефективним для ідентифіка-ції породної належності бджіл обох порід. Ліворуч під умовною назвою Apis mellifera acervorum показано відтворення ДНК-фінгерпринтів зразків бджіл української породи, праворуч — карпатської (Apis mellifera carpatica).
Як свідчить фотовідбиток, ДНК-маркери, позначені символами Са1, Са2, Са3, Са4, виявлені в усіх фінгерпринтах бджіл карпатської і були відсутні у бджіл української породи. Ці ДНК-маркери чітко розрізняють досліджуваних бджіл і є придатними для ідентифікації бджолиних сімей української і карпатської порід, проявляючи добру відтворність ДНК-фінгерпринтів.
Важливою особливістю результатів ДНК-тестування бджіл, як видно з рисунка, є висока генетична однорідність досліджуваних сімей в межах породи.
Порівняння результатів досліджень на генетичну дактилоскопію показало не тільки добре виражені породні відмінності українських і карпатських бджіл, а й відсутність у них ознак сірої гірської кавказької і середньоросійської порід, досліджених раніше в лабораторії.
RAPD-ампфлікації з праймером В15 в 6% ПААГ зразків бджіл української і карпатської порід
Встановлена в наших дослідах видова специфічність генома бджіл, відібраних у типових умовах ареалу їх існування (Карпати і Лісостеп України), дає підставу стверджувати про належність карпатських та українських бджіл до різних порід, які характеризуються своїми особливостями за ознаками екстер’єру, поведінки, пристосованості до збирання корму, стійкості проти впливу факторів середовища.
Отримані нами дані щодо характеристики досліджуваних порід вказують на необхідність аналізу результатів з вивчення історії їх формування, методів розведення, селекції, районування при сучасному господарському використанні та збереження у чистоті в своєму природному ареалі.
Привертає увагу, зокрема, той факт, що генетична однорідність бджіл хмельницького внутрішньопородного типу, який складається з дев’яти генеалогічних груп, збігається з ознаками консолідованості за результатами вивчення промірів екстер’єру. Так, міжгрупові коливання середньої довжини хоботка становлять 6,34-6,63 мм, кубітального індексу — 2,18-2,62, дискоїдального зміщення — 72-94% позитивних випадків, випукла форма заднього краю воскового дзеркальця п’ятого стерніта — 64-96%.
Селекція бджіл методом чистопородного розведення з одночасним розподілом відібраного матеріалу на генеалогічні групи з використанням основних породних ознак екстер’єру для постійного контролю належності сімей до української породи підтверджена методом виявлення ДНК-маркерів.
Відсутність у них ознак інших порід вказує на можливість утримання чистопородних сімей в осередках напівзакритої популяції і достатню ефективність їх використання та контролю якості материнської групи за ознаками екстер’єру бджіл.
Висновки. Проведений аналіз показав, що бджоли української породи за результатами ДНК-тестування відрізняються від бджіл гірських районів Карпат, їм властиві свої генетичні відмінності.
У межах породи досліджувані бджоли характеризуються високою однорідністю і добре диференціюються за допомогою встановлених ДНК-маркерів.
Список літератури
- Аветисян Г. А., Давиденко І. К., Макаров Ю. І. Досвід племінної роботи з карпатськими та далекосхідними бджолами // Бджільництво. — К.: Урожай. — 1969. — Вип. 5. — С. 3-7.
- Билаш Г. Д., Кривцов Н. И. Селекция пчел. — М.: Агропромиздат, 1991. — 302с.
- Гайдар В., Пилипенко В. 50 лет на кафедре пчеловодства в ТСХА // Пчеловодство. — 1996. — № 3. — С. 2.
- Гайдар В. А., Левченко И. А. Сравнительная оценка карпатских и краинских пчел. // Пчеловодство. — 2003. — № 8. — С. 18-21.
- Глазко В. И., Глазко Г. В. Словарь терминов по прикладной генетике и ДНК-технологиям. — К.: Нора-принт, 1999. — 342 с.
- Глазко В. И., Доманский Н. Н., Созинов А. А. Современные направления использования ДНК-технологий // Цитология и генетика. — 1998. — Т. 32. — № 5. — С. 80-93.
- Капралова О. В., Дубова Л. В. Содержание ДНК — критерий породы пчел // Пчеловодство. — 1989. — № 3. — С. 14-15.
- Нестерводський В. А. Організація пасік і догляд за бджолами. — К.: Урожай. — 1966. — 396 с.
- Нестерводський В. А. Як одержати більше меду та воску. — К.: Держсільгоспвидав УРСР, 1960. — 76 с.
- Нормативно-правові акти з питань розвитку бджільництва. — К.: Мінагрополітики України, 2001. — 103 с.
- Поздняков В. Н., Абрамова А. Б., Чудинов О. С., Какпаков В. Т., Козин Р. Б., Кравцов Н. И., Бородачев А. В. Молекулярно-генетические подходы в изучении генетического полиморфизма различных пород пчел // Сельскохозяйственная биология. — 2000. — № 4. — С. 56-59.
- John G., Williams K. e. a. Genetic analysis using RAPD-markers. Methods in Enzymology, 1993. — 218 р.
- Williams J., Kubelik A., Livak K., Rafalski K., Tingey J. DNA polimorphism amplified by arbitrary primers are useful as genetic markers // Nucleic Acids Rec. — 1990. — Vol. 18. — P. 6513-6535













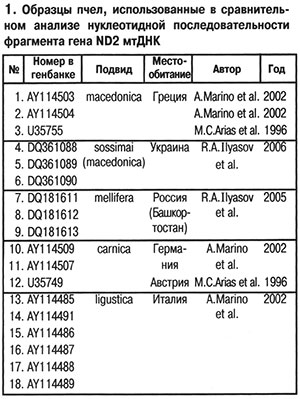 Для уточнения действительного таксономического статуса украинских пчел необходимо было провести дополнительные исследования с использованием молекулярно-генетических методов. С этой целью мы сравнили последовательности нуклеотидов во фрагменте гена ND2 митохондриальной ДНК (мтДНК) у ряда подвидов европейских пчел. Исследование провели на трех образцах пчел с Украины (пасека А.Д.Комиссара около Киева), трех образцов из России (Республика Башкортостан, Бурзянский район: пасеки «Капова Пещера», «Коран-Елга», борти Государственного природного заповедника «Шульган-Таш») и внесли в базы данных международных генетических банков (табл. 1).
Для уточнения действительного таксономического статуса украинских пчел необходимо было провести дополнительные исследования с использованием молекулярно-генетических методов. С этой целью мы сравнили последовательности нуклеотидов во фрагменте гена ND2 митохондриальной ДНК (мтДНК) у ряда подвидов европейских пчел. Исследование провели на трех образцах пчел с Украины (пасека А.Д.Комиссара около Киева), трех образцов из России (Республика Башкортостан, Бурзянский район: пасеки «Капова Пещера», «Коран-Елга», борти Государственного природного заповедника «Шульган-Таш») и внесли в базы данных международных генетических банков (табл. 1).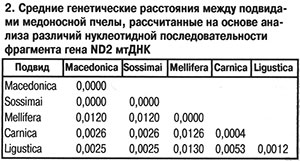 На основании сравнения нуклеотидных последовательностей фрагмента гена ND2 мтДНК пчел с использованием программы MEGA 3.1 (Kumar et al., 2004) и дальнейшего расчета средних генетических расстояний между образцами было показано тесное генетическое родство пчел с Украины (пасека А.Д.Комиссара) и пчел A. m. macedonica из Греции: средние генетические расстояния между ними были равны 0,0000 (табл. 2). Наиболее генетически отдаленными от украинских пчел оказались среднерусские A. m. mellifera из Республики Башкортостан (Россия), что вполне логично, так как A. m. macedonica, A. m. carnica, A. m. ligustica — представители эволюционной ветви C (Ruttner, 1988), а A. m. mellifera — эволюционной ветви М.
На основании сравнения нуклеотидных последовательностей фрагмента гена ND2 мтДНК пчел с использованием программы MEGA 3.1 (Kumar et al., 2004) и дальнейшего расчета средних генетических расстояний между образцами было показано тесное генетическое родство пчел с Украины (пасека А.Д.Комиссара) и пчел A. m. macedonica из Греции: средние генетические расстояния между ними были равны 0,0000 (табл. 2). Наиболее генетически отдаленными от украинских пчел оказались среднерусские A. m. mellifera из Республики Башкортостан (Россия), что вполне логично, так как A. m. macedonica, A. m. carnica, A. m. ligustica — представители эволюционной ветви C (Ruttner, 1988), а A. m. mellifera — эволюционной ветви М.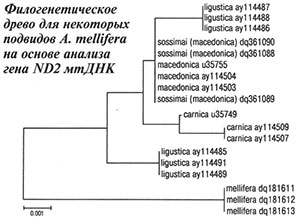 По данным генетических дистанций методом кластеризации ближайшего соседа (Saitou, Nei, 1987) мы построили филогенетическую схему (рис.). На ней видно, что все образцы A. m. sossimai и A. m. macedonica объединились в одну группу, что говорит о тесном генетическом родстве украинских и греческих пчел. В другую группу объединились пчелы A. m. carnica из Германии и Австрии, которые из всех сравниваемых подвидов были наиболее близки к группе A. m. macedonica. Представители A. m. mellifera из Республики Башкортостан оказались наиболее отдаленными от группы, объединяющей A. m. macedonica и A. m. sossimai, а также от групп других подвидов эволюционной ветви С. Образцы пчел A. m. ligustica из Италии разделились на две группы, одна из которых была близка к группе A. m. macedonica, а другая находилась на значительном отдалении от A. m. macedonica и A. m. carnica и была близка к A. m. mellifera. Согласно результатам исследований морфометрических характеристик пчелы из Италии обладали бóльшим сходством с A. m. carnica из Австрии и Югославии (Ruttner, 1988). Возможно, некоторые популяции A. m. ligustica из Италии пересекались с пчелами A. m. mellifera из Франции, что подтверждает полиморфизм межгенного локуса COI-COII мтДНК (Franck et al., 2000).
По данным генетических дистанций методом кластеризации ближайшего соседа (Saitou, Nei, 1987) мы построили филогенетическую схему (рис.). На ней видно, что все образцы A. m. sossimai и A. m. macedonica объединились в одну группу, что говорит о тесном генетическом родстве украинских и греческих пчел. В другую группу объединились пчелы A. m. carnica из Германии и Австрии, которые из всех сравниваемых подвидов были наиболее близки к группе A. m. macedonica. Представители A. m. mellifera из Республики Башкортостан оказались наиболее отдаленными от группы, объединяющей A. m. macedonica и A. m. sossimai, а также от групп других подвидов эволюционной ветви С. Образцы пчел A. m. ligustica из Италии разделились на две группы, одна из которых была близка к группе A. m. macedonica, а другая находилась на значительном отдалении от A. m. macedonica и A. m. carnica и была близка к A. m. mellifera. Согласно результатам исследований морфометрических характеристик пчелы из Италии обладали бóльшим сходством с A. m. carnica из Австрии и Югославии (Ruttner, 1988). Возможно, некоторые популяции A. m. ligustica из Италии пересекались с пчелами A. m. mellifera из Франции, что подтверждает полиморфизм межгенного локуса COI-COII мтДНК (Franck et al., 2000).